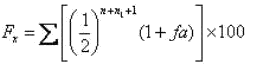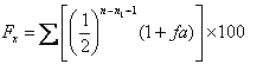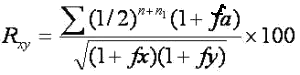|
КАЛЕНДАРНЫЙ МОДУЛЬ 1
ДИСЦИПЛИНАРНЫЙ МОДУЛЬ 6
Тема 6.2 Родственное спаривание
6.2.1 Понятие об инбридинге. Отношение к инбридингу в различное время
6.2.2 Учет степени инбридинга
6.2.3 Вредные последствия близкородственных спариваний и меры их предупреждения
6.2.4 Гетерозис и его использование в животноводстве
Вопросы для самоконтроля
6.2.1 Понятие об инбридинге. Отношение к инбридингу в различное время
Инбридингом (родственным спариванием, инцухтом) принято считать такое спаривание, п ри котором отец и мать будущего потомства состоят между собой в кровном родстве, имеют одного или несколько общих предков. В противоположность инбридингу различают аутбридинг, т. е. неродственное спаривание.
Обычно случаи применения инбридинга устанавливают по родословным животных. Если в родословной со стороны матери и отца в пределах первых пяти рядов предков встречается одно и то же животное (или несколько животных), то считают, что данное животное (пробанд) получено путем применения инбридинга. Если в материнской и отцовской частях родословной пробанда в пяти рядах нет общих предков или они встречаются с учетом пятого и более далеких рядов предков, то пробанд является аутбредным.
Инбридинг представляет собой крайнюю форму однородного (гомогенного) подбора животных по происхождению. Он может быть простым (на одного предка) и сложным, или комплексным (на двух и более предков). Для быстрого определения инбредности пробанда его предков, на которых применялся инбридинг, помечают в родословной каким-либо общим знаком. В различные исторические времена человек по-разному относился к инбридингу.
С глубокой древности у людей выработалось отрицательное отношение к родственному спариванию. Это, скорее всего, было следствием того, что в процессе приручения и одомашнивания вынужденно приходилось спаривать между собой животных близких степеней родства.
Затем Шапоруж (1909) показал, что инбридинг умеренных степеней давал и хорошие результаты и применялся даже в те времена, когда наблюдалась общая боязнь родственных спариваний.
В конце XIX века П.Н. Кулешов (1890) открыто выступил с защитой инбридинга как важного приема племенной работы. Основываясь на практике тонкорунного овцеводства, он пришел к выводу, что инбридинг имеет большое значение в совершенствовании разных пород животных.
Е.А. Богданов (1916) впервые высказал предположение, что разные степени родства по-разному влияют на результат применения инбридинга, связывая это с возрастанием гомозиготности.
Большой вклад в разработку теоретических основ инбридинга и практических приемов его использования в животноводстве внесли такие известные ученые, как С. Райт, Д. А. Кисловский, М. Ф. Иванов и другие.
Чтобы разобраться в природе и последствиях инбридинга, необходимо рассмотреть вопрос о том, как он учитывается и какие степени различают.
6.2.2 Учет степени инбридинга
Для измерения степени инбридинга в разное время были предложены различные способы. Некоторые из них не нашли применения в практике и представляют лишь чисто исторический интерес, являются иллюстрацией различных подходов к определению степени родства.
В настоящее время для учета степени инбридинга широко используют способы, предложенные Шапоружем (1909), Райтом (1921) и Кисловским (1925).
Способ Шапоружа заключается в том, что ряды предков пробанда в родословной, начиная с родительского, обозначают римскими цифрами. Затем записывают ряд, в котором общий предок встречается сначала в материнской части родословной, а потом - в отцовской. Если предок в одной стороне родословной повторяется несколько раз, то записывают все ряды, в которых он встречается, разделяя их запятыми, а если в другой отсутствует, то со стороны, где его нет, ставят ноль. При этом считают, что пробанд получен при спаривании неродственных животных, но один из его предков был инбридированным.
Так, корова ярославской породы Гамма 1907 (табл. 1) получена в результате применения комплексного инбридинга в степени II-III на Невода 509.
Исходя из этой системы записи, в настоящее время различают следующие степени инбридинга: кровосмешение (очень тесный) (I-II‚ II-I‚ II-II), близкое родственное спаривание (тесный инбридинг) (I- III, III-I, II-III‚ III-II) и умеренное родственное спаривание (III-III, III-IV, IV- III, IV-IV). Более далекие степени родственных спариваний относятся к отдаленному инбридингу, который, как правило, не учитывается. Самым тесным кровосмешением является спаривание однопометных животных. Спаривание матери с сыном считается более тесным, чем отца с дочерью. Это связано с двойным влиянием материнского организма на потомство. Использование способа учета степеней инбридинга по Шапоружу, особенно при комплексном инбридинге, часто не дает возможности установить, в каком случае животные были больше инбридированы, т.е. не позволяет количественно выразить степень инбридинга. Не отражает этот способ и генетических последствий инбридинга, которые заключаются в том, что он ведет к уменьшению гетерозиготности и возрастанию гомозиготности.
Таблица 1 - Гамма 1907 ЯЯ-17312, ярославской породы, 3-252-3474-5,7
| Жадовка 97, 4-300-3806-4,9 эл. |
Великан 487 эл.-р. |
| Невруша 593 8-300-5959-4,39 эл.-р |
Невод эл.-р. 509 ♦ |
Недоучка 577 5-295-4357-4,2 эл. |
Атрибут 1268 эл.-р |
| Груша 302 7-300-4350-4,2 эл.▲ |
Быстрый 270 эл.-р. |
Дина 6 7-300-4116-5,08 эл.-р. |
Баян 14 эл.-р. |
Дочурка 246 3-300-3112-4,81 |
Вымпел 61 эл. |
Груша 302 7-300-4350-4,2 эл. ▲ |
Невод эл.-р. 509 ♦ |
С. Райт (1921) предложил вычислять в качестве меры инбридинга коэффициент инбридинга, или коэффициент возрастания гомозиготности. С этой целью обычно используют формулу С. Райта:
где Fх - коэффициент инбридинга;
∑ - знак суммирования в случае комплексного инбридинга;
½- доля наследственности, получаемой пробандом от каждого предка в зависимости от того, в каком ряду родословной он находится;
n - ряд предков, в котором общий предок встречается в материнской части родословной;
n1 - ряд предков, в котором общий предок встречается в отцовской части родословной;
fa - коэффициент инбридинга общего предка, если он инбридирован (при этом счет рядов предков ведется не с родительского, а с дедовского ряда).
Коэффициент инбридинга выражается в долях единицы или в процентах и может быть в пределах от 0 до 1 или от 0 до 100%. Он показывает не абсолютную гомозиготность инбридированных особей, а лишь вероятную степень ее возрастания у них по сравнению с животными, полученными при аутбредных спариваниях.
Коэффициент инбридинга - это среднестатистическая величина. Если он, допустим, равен 25%, то это не означает, что у данного животного гомозиготность возрастет точно на 25%. Данная величина показывает, что при получении многих животных с использованием таких же степеней родственных спариваний гомозиготность у них в среднем возрастет на 25%.
Д.А. Кисловский в формулу С. Райта внес изменения и преобразовал ее:
где Fх - коэффициент инбридинга, выраженный в процентах;
∑ - знак суммирования в случае комплексного инбридинга;
½- доля наследственности, получаемой пробандом от каждого предка в зависимости от того, в каком ряду родословной он находится;
n, n1 - ряды в материнской и отцовской сторонах родословной, где встречается общий предок, но в отличие от формулы С. Райта счет рядов предков начинается с родительского ряда.
По Д.А. Кисловскому, при коэффициенте 25% и более инбридинг считается тесным (кровосмешение), от 12,5 до 25% - близким, от 1,55 до 12,5% - умеренным, от 0,20 до 1,55% - отдаленным.
Для измерения генетического сходства между животными С. Райт предложил следующую формулу:
где Rxy - коэффициент генетического сходства между животными х и у, %;
n - ряд в родословной животного х, в котором встречается общий предок (по Шапоружу);
n1 - ряд родословной животного у, в котором встречается общий предок;
fа - коэффициент инбридинга (по Кисловскому) для общего предка (в десятичных дробях);
fy - коэффициент инбридинга (по Кисловскому) для животного у (в десятичных дробях);
fx - коэффициент инбридинга (по Кисловскому) для животного х (в десятичных дробях).
Вычисление коэффициента генетического сходства аналогично вычислению коэффициента инбридинга.
У коровы Гамма 1907, родословная которой приведена выше, коэффициент инбридинга составляет 6,25%.
6.2.3 Вредные последствия близкородственных спариваний и меры их предупреждения
Анализ многочисленных материалов показывает, что степень вреда от родственного спаривания связана со степенью родства спариваемых животных. Чем ближе инбридинг, тем ярче выражены его отрицательные последствия.
Инбридинг и в первую очередь его самые тесные степени - самоопыление у растений, кровосмешение и близкое родство у животных - сопровождается снижением жизнеспособности и уровня развития различных признаков, которое получило название инбредной депрессии. Ее обычно наблюдают в тех случаях, когда у потомков определенные признаки имеют меньшую величину, чем у худшей родительской формы. Инбредная депрессия особенно сильно проявляется в первых поколениях инбридированных особей и снижается в последующем до тех пор, пока не достигнет стабильной величины - инбредного минимума. Она в наибольшей степени затрагивает те признаки, которые связаны с приспособляемостью животных (плодовитость, материнские качества, выживаемость, скорость роста и т. д.). Обычно эти признаки характеризуются низкими коэффициентами наследуемости.
Вредные последствия инбридинга обусловлены изменением морфофизиологических и биохимических особенностей организма животного, таких, как ферментативная активность, способность переваривать и использовать питательные вещества корма, резистентность и др.
В обобщенном виде вредные последствия близких степеней инбридинга в животноводстве можно свести к следующему.
1. Происходит снижение плодовитости животных, в крайних случаях - до полного бесплодия. В этом можно убедиться при анализе данных таблицы 2.
Таблица 2 - Снижение плодовитости у инбредных кур (по М.М. Кот, 1990)
Поколение от
спаривания типа
брат × сестра |
Число инку-
бируемых яиц |
Доля оплодот-
воренных яиц, % |
Доля здоровых цыплят, % |
от оплодот-
воренных яиц |
от заложенных яиц |
| 1 |
5452 |
90,7 |
68,6 |
62,4 |
| 2 |
1811 |
88,0 |
62,4 |
54,9 |
| 3 |
3768 |
86,0 |
53,3 |
45,9 |
| 4 |
1503 |
64,0 |
43,1 |
32,1 |
У кур четвертого поколения по сравнению с первым оплодотворенность яиц снизилась на 26,7%, доля полученных здоровых цыплят от оплодотворенных яиц уменьшилась на 25,5%.
2. Снижается жизнеспособность, наблюдаются общее ослабление конституции животных, переутончение скелета, изнеженность, животные легко подвергаются различным неблагоприятным внешним воздействиям.
М.Ф. Иванов (1949) отмечает, что в процессе выведения украинской степной белой породы свиней при разведении полукровных животных (местная украинская?крупная белая) "в себе" с применением родственных спариваний отход молодняка составил 10,2% в первом поколении и 13,7% - во втором. Среди полукровных животных и животных, полученных в результате скрещивания помесей первого поколения с хряками крупной белой породы, отхода не было.
3. Ухудшается развитие животных, происходит измельчение потомства. Об этом свидетельствуют данные таблицы 3.
Таблица 3 - Влияние инбридинга на живую массу и воспроизводительную функцию коров голштинской породы (по М.М. Кот, 1990)
Поколение
от тесного
инбридинга |
Коэффициент
инбридинга, % |
Живая масса, кг |
Число
спариваний
на одно
оплодотворение |
| при рождении |
в 24 мес. |
| I |
0 |
39 |
505 |
2,18 |
| II |
26 |
35 |
446 |
2,38 |
| III |
33 |
34 |
438 |
2,78 |
| IV |
44 |
32 |
432 |
2,81 |
| V |
45 |
30 |
433 |
3,34 |
| VI, VII |
60 |
26 |
402 |
4,57 |
4. Снижается продуктивность. Хотя в отдельных случаях, как видно из приведенных данных (табл. 4), инбридинг позволяет получать высокопродуктивных животных, все же в массе он ухудшает показатели продуктивности, особенно при тесных степенях инбридинга и длительном его применении.
Таблица 4 - Молочная продуктивность инбредных и аутбредных коров швицкой породы по I лактации (по М.М. Кот, 1990)
Тип
подбора |
Количество,
гол. |
Живая
масса, кг |
Удой, кг |
Содержание
жира, % |
| Кровосмешение |
20 |
478±7,5 |
3130±223 |
3,76±0,03 |
Близкое
родство |
164 |
482±2,9 |
3110±73 |
3,81±0,01 |
Умеренное
родство |
204 |
489±2,3 |
3427±60 |
3,80±0,01 |
Отдаленное
родство |
228 |
491±2,3 |
3361±60 |
3,79±0,01 |
| Аутбридинг |
741 |
487±1,3 |
3269±33 |
3,80±0,00 |
5. Могут появляться уроды. При запущенном зоотехническом учете и стихийном инбридинге в свиноводстве можно часто наблюдать появление животных с различными отклонениями от нормального развития: с аномальной головой, недоразвитыми конечностями, отсутствием анального отверстия и др. У крупного рогатого скота описано более 10 различных уродливых форм, появляющихся при применении инбридинга. Известны различные уродства у овец, коз, кроликов и других видов животных.
Инбридинг может применяться только в племенных хозяйствах при наличии высококвалифицированных специалистов. Чтобы не допустить автоматического, не вызываемого зоотехнической необходимостью инбридинга, раньше между хозяйствами практиковался периодический обмен производителями. В настоящее время в молочном скотоводстве применяется чередование в использовании быков-производителей, принадлежащих к разным линиям: через каждые два года используются в хозяйстве производители другой линии, что по идее должно автоматически исключать родственные спаривания. В связи со злоупотреблениями кроссами при разведении животных по линиям этот прием часто не дает желаемых результатов, не позволяет избегать инбридинга. Так, С.И. Харитонов (по Л.К. Эрнсту, 1986) показал, что в Московской области, несмотря на ротацию линий, удельный вес инбредных коров составил 29,35%, в том числе по 3,07% приходится на коров, полученных при кровосмешении и при близком родственном спаривании. У бабушек всех обследованных коров (ММ) он составлял лишь 11,51%, т.е. за два поколения увеличился в 2,55 раза. Такие тревожные данные послужили основанием для рекомендаций специализировать каждый племзавод на работе только с одной линией.
Если инбридинг уже имел место и привел к отрицательным последствиям, необходимо провести освежение крови животных. Суть этого приема заключается в том, что в стаде используют производителей той же породы, желательно сходных по типу с животными стада, но неродственных им, полученных в других стадах или регионах.
При применении инбридинга животным следует создать благоприятные условия кормления и содержания. В неудовлетворительных условиях использования животных инбридинг может принести только вред.
Лучшие результаты получают в тех случаях, когда предназначенные для родственных спариваний животные воспитываются в несходных, различающихся условиях кормления и содержания.
Инбридинг необходимо применять на конституционально крепких и через конституционально крепких животных. Наличие конституциональных различий между спариваемыми родственниками обычно способствует ослаблению инбредной депрессии у потомков.
Если у животных в результате применения инбридинга появились какие-то отрицательные признаки, то никакими мерами их не исправить. Самым радикальным выходом из ситуации являются отказ от родственных спариваний, выбраковка всех, не удовлетворяющих предъявляемым требованиям животных и использование аутбридинга.
6.2.4 Гетерозис и его использование в животноводстве
Термин "гетерозис" (в переводе с греческого языка - изменение, превращение) ввел в 1914 году американский исследователь А. Шелл вместо термина "гетерозигозис", которым по предложению американского ученого Е. Иста обозначали "гибридную силу" с 1907 года.
Под гетерозисом обычно понимают свойство помесных животных или гибридов первого поколения превосходить по конкретным признакам лучшую из родительских форм.
В животноводческой практике это явление было известно и использовалось с древних времен: производство мулов, т.е гибридов между лошадью и ослом, у которых ярко выражен гетерозис по крепости конституции, жизнеспособности, выносливости и долголетию, практикуется более 2000 лет.
Гетерозис - явление сложное, он свойствен не всем признакам в одинаковой степени. Как правило, гетерозис проявляется по тем признакам, которые больше подвержены инбредной депрессии и характеризуются невысокой наследуемостью. Наиболее выражен гетерозис по признакам, развивающимся у животных в ранний период жизни (например, выживаемость, скорость роста до отъема и др.). В меньшей степени он проявляется по таким признакам, как скорость и эффективность роста после отъема и др., которые формируются у животных в более поздние периоды индивидуального развития.
Проявление гетерозиса по различным хозяйственно полезным признакам имеет свои особенности. По признакам, которые подвергались очень длительной селекции (молочная продуктивность у коров, резвость у лошадей), обычно лучшие результаты наблюдаются не у помесей первого поколения, а у чистопородных животных или помесей с более высокой кровностью по одной из пород. При этом большое значение имеет сочетаемость линий при чистопородном разведении животных.
В животноводстве относительно больший эффект гетерозиса получают при спаривании животных разных линий из разных пород, особенно если они существенно различаются между собой.
Гетерозис, как принято считать, зависит от неаддитивного действия генов - доминирования, сверхдоминирования и эпистаза - и от гомозиготности родителей по разным генам одного и того же локуса (например, АА и аа, СС и сс и т. д.).
Все коровы - рекордистки по молочной продуктивности - не являются помесями первого поколения. Им свойственны, вероятно, не просто высокая гетерозиготность, а благоприятная комбинация, выщепление каких-то полезных генов, что является результатом длительного отбора. Иммуногенетические исследования показали, что высокие удои бывают у животных при примерно равном отношении локусов в гомо- и гетерозиготном состоянии. Отсюда следует, что какие-то гены обязательно должны быть в гомозиготном состоянии и организм в целом отличается высокой жизнеспособностью, которая является следствием его гетерозиготности по части локусов.
В литературе имеются данные о том, что эффект гетерозиса зависит от гомо- или гетерогаметности пола. У животных, характеризующихся гомогаметным полом, он выше. Известно, что у млекопитающих гомогаметным полом являются самки, а гетерогаметным - самец; у птиц, наоборот, к гомогаметному полу относятся самцы.
Встречаются разные классификации форм проявления гетерозиса. Т. Добжанский (1950) подразделяет гетерозис на три формы: 1 - приспособительная, характеризующаяся тем, что особи способны адаптироваться и приспосабливаться даже к весьма неблагоприятным условиям внешней среды; 2 - пышного развития, при котором наблюдается повышенная скорость роста, особи характеризуются большей массой; 3 - мутационная, связанная с освобождением инбредной линии от вредных рецессивных генов, элиминация которых в исходной популяции затруднена, так как они прикрыты доминантными генами и фенотипически невозможно определить, является особь их носителем или нет.
Густавсон (1951) также выделяет три формы гетерозиса: 1 - репродуктивная, при которой наблюдаются повышенная плодовитость и многоплодие; 2 - соматическая (аналогичная форме пышного развития); 3 - приспособительная.
Иногда все эти формы гетерозиса проявляются вместе, чаще каждая из них встречается отдельно.
На основе обобщения обширного материала о проявлении гетерозиса у сельскохозяйственных животных X. Ф. Кушнер выделяет следующие его формы:
1 - помеси и гибриды первого поколения превосходят своих родителей по живой массе и жизнеспособности. Эта форма наблюдается при межпородном скрещивании в свиноводстве, мясном скотоводстве, при спаривании одногорбых и двугорбых верблюдов;
2 - помеси первого поколения по живой массе занимают промежуточное положение между родителями, но заметно превосходят их по плодовитости и жизнеспособности. Иллюстрацией такого проявления гетерозиса могут служить потомки от скрещивания кур породы белый леггорн с породами нью-гемпшир, плимутрок, австралорп и другими;
3 - гибриды первого поколения превосходят исходные формы по конституциональной крепости, долголетию, физической работоспособности, но полностью или частично теряют плодовитость. Мулы, например, в отличие от родителей, имеют очень крепкую конституцию, промежуточную живую массу и практически полностью бесплодны. При гибридизации крупного рогатого скота с яками, бизонами, гаялами, а также зебу с яками плодовитыми являются только самки. Гибридные самцы бесплодны не только в первом, но и во втором, а часто и в третьем поколениях от обратных скрещиваний. При отдаленной гибридизации птиц нормальное развитие чаще всего отмечается только у особей мужского пола. Самки обычно погибают в эмбриональный период своего развития или вскоре после вылупления.
4 - у помесей или гибридов каждый отдельно взятый признак наследуется промежуточно, а в отношении конечной продукции, являющейся производным этих признаков, наблюдается типичный гетерозис. Например, при скрещивании черно-пестрой и джерсейской пород скота удой и содержание жира в молоке первого поколения средние, а выход молочного жира в удое выше, чем у родителей (табл. 5).
Таблица 5 - Гетерозис по содержанию молочного жира в удое коров при скрещивании голштинской и джерсейской пород
Группа
животных |
Удой по
1-й лактации,
кг |
Содержание
жира, % |
Продукция
молочного
жира, кг |
| Голштинская порода |
6537 |
3,59 |
233 |
| Джерсейская порода |
3718 |
5,46 |
201 |
Помеси F1 от
голштинских коров
и джерсейских быков |
5588 |
4,61 |
255 |
Помеси F1 от
реципрокного
скрещивания |
5808 |
4,88 |
282 |
5 - помеси не превосходят по признакам лучшую из родительских форм, у них признаки лишь выше, чем в среднем у родителей, что отмечалось выше.
Приведенные формы проявления гетерозиса касаются хозяйственно полезных признаков животных. Различные интерьерные показатели также неодинаково изменяются: одни наследуются промежуточно, по другим помеси превосходят в разной степени родительские формы, т.е. какого-то проявления гетерозиса нет. Формы его исключительно многообразны.
Вопросы для самоконтроля
- Какое спаривание называют родственным?
- Как определить степень родственного спаривания по Шапоружу?
- Как рассчитывают коэффициент инбридинга по Райту и Кисловскому?
- Каковы биологические последствия различий степеней родственного спаривания?
- Что такое явление гетерозиса? Пути его получения.
- Что такое генетическое сходство, как его вычислить?
- В чем сущность инбредной депрессии?
|